Answers
In 1955, Vernon Mountcastle discovered the first evidence for functional columnar organization in the somatosensory cortex of the cat. Hubel and Wiesel reported similar evidence for columnar structure in their 1962 paper on the visual cortex of the cat. Additional research has uncovered other kinds of columnar organization, including so-called "ocular dominance columns".
Despite the importance of the column hypothesis for inspiring fundamental work in neuroscience, contemporary experimental methods and techniques are calling into question the idea that the brain is organized as a chain of processing units, each composed of an "ice-cube tray" of independent, columnar computational elements.
Mountcastle and de Nó: Columns in Somatosensation and Development
In a series of ground-breaking experiments, Vernon Mountcastle recorded the responses of somatosensory cortical neurons in the anesthetized cat to light mechanical stimulation and joint rotation using extracellular electrodes.

Vernon Mountcastle, quite the cheery fellow. From azQuotes.com.
These experiments confirmed the existence of a somatotopic map in the cortex. X-topic maps refer, in neuroscience, to regions where smooth movement on the surface of the brain causes smooth movement in some stimulus space X. In this case, somatotopic means that the stimulus space is the soma (Greek, "body") of the cat, and so moving our electrode across the surface of the brain causes the spot of the cat's body where mechanical manipulation evokes a neural response to move.
More critically for the purposes of this question, Mountcastle found that neurons of the somatosensory cortex responded to either soft touching or to joint rotation, but never to both.
Even more intriguingly, he found that if he moved his electrode up or down (that is, towards the surface or towards the interior), the neurons in this new location still responded to the same kind of stimulation in the same spot. If he moved his electrode to the side, however, the neurons in this lateral location would respond to either a different kind of stimulation, stimulation in a different spot, or both.
Since the neurons were stacked on top of each other, Mountcastle called these functional units "columns". Part of the excitement around this finding was that, anatomically speaking, it seemed extremely reasonable.
First, going back to the earliest days of neuroscience, Ramón y Cajal had recognized a certain vertical regularity to the cortex (see the drawing below).
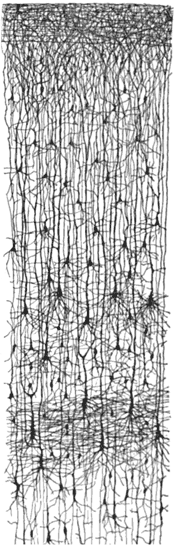
This drawing shows a collection of cortical pyramdial neurons (so named because of their triangular shape) The neurons' axons are aligned, all reaching towards the cortical surface (top of the image). The relative paucity of horizontal connections had led many to hypothesize that cortex was vertically oriented.
This idea was shored up by some findings from developmental neuroscience. As discovered by Lorento de Nó in 1949, pyramidal neurons are born from precursor cells at the bottom of cortex, and then travel upwards to their final position. Unlike many other cells in the body, which move in complex patterns as they squeeze into laminae or clump into organs, the neurons moved in straight lines, perpendicular to the surface.
This meant that neurons lying along a column were genetically "siblings", all born from the same progenitor cell. This genetic similarity would suggest a functional similarity, as de Nó proposed and Mountcastle seemed to confirm.
Hubel and Wiesel: Columns and Hypercolumns in Visual Cortex
In addition to discovering and describing simple cells and complex cells, the ground-breaking, Nobel-winning experiments of David Hubel and Torsten Wiesel also provided additional support for the column hypothesis.
They similarly found that receptive fields varied greatly when they moved laterally, but remained similar when they moved vertically. However, their superior characterization of the properties of these fields allowed them to more deeply describe changes and invariances as they moved in both directions.
The cells identified by Hubel and Wiesel were orientation-selective edge detectors of varying kinds. As they moved their electrode laterally, the preferred orientation changed. If they moved their electrode up and down, the preferred orientation stayed the same, but the other characteristics of the receptive field would change: the first neuron might prefer moving, bright bars, while the next above it would prefer stationary, dark bars, and then above that a neuron that preferred much shorter or longer bars than either of the two previous neurons.
If you've never seen them before, check out this video from a TV documentary or this video from an actual experiment.
In addition to this form of anatomical organization, visual cortex also has a -topic map, just like the somatotopic map in somatosensory cortex. As electrodes move laterally over long distances, the location of the receptive field moves. That is, light hitting the retina in a different location causes the cells to activate. This kind of map is retinotopic.
Combining these pieces of information, Hubel and Wiesel theorized the existence of "hypercolumns", or collections of columns, all sharing the same retinal coordinate but with different receptive fields. With this setup, each point in the visual field will be subjected to analysis by every kind of receptive field at every orientation.
To make this picture more clear, let's imagine what this organization of cortex would look like. The image below shows this organization for a chunk of (fictitious) emoji cortex, recorded from the brain of an anesthetized millennial†.
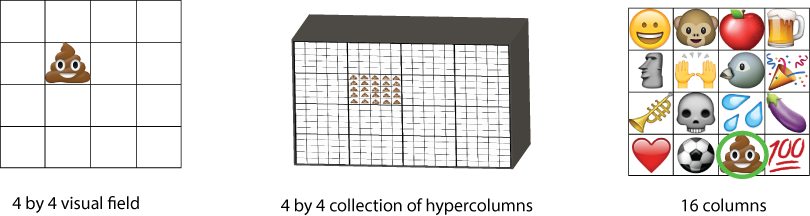
On the left, we have the visual field, which is particularly simple: just 16 possible locations, arranged in a square. An emoji appears in the sixth position.
This stimulus is processed by the emoji cortex, center. There are 16 hypercolumns, arranged retinotopically – the top left hypercolumn corresponds to the top left area of the visual field, and so on. The surface is facing us, and columns run into the screen. Every column within the hypercolumn gets a copy of the stimulus.
The right panel zooms in on 16 of these columns. The neurons in each column share a receptive field property: they each respond to one particular emoji, the skull, the aubergine, the praisemoji, etc. Within a column, the neurons have varying additional properties. Perhaps different neurons detect the emoji under different lighting conditions, or from different angles, or specialize in the Apple version or the Google version.
The result is that some neurons in the column circled in green become active, since the stimulus matches their preferred stimulus (pile-of-poo).
In the Hubel and Wiesel model, the emojis would be replaced by oriented bars, with columns specializing in different orientations, rather than different emojis.
More Columns: Ocular Dominance
Hubel and Wiesel discovered another form of columnar organization in the macaque cortex, which they reported in a 1969 paper.
Animals with high-quality stereoscopic vision (generally, predators or primates) use the disparity, or difference, between the 2D images presented to their two eyes to compute the 3D structure of the world. This means that the two parallel streams of information, one from each eye, must be combined at some point.
In the early visual system of the macaque, they are kept mostly separate. In fact, the two inputs are organized into columns, resulting in columns of neurons with the same receptive field location, orientation tuning, and dominant eye.
A Coda on Columns
Much of the information in this blog post came from a review of the cortical column literature published by Horton and Adams on the 50th anniversary of Mountcastle's discovery. The title was "The Cortical Column: A Structure Without a Function", which indicates the unfortunate state of research into columnar organization and cortical microcircuitry.
Many of the classic findings described above have been challenged. For example, the connection between ontogenetic or developmental columns and functional columns remains unclear, though recent work from Yang Dan's lab shows that developmentally-related neurons in mouse visual cortex are functionally similar. Note, however, that unlike macaques and cats, mice lack orientation columns.
In fact, many mammals lack columnar structure in their visual cortices. It was long hypothesized, based on the classic findings from mice, cats, and monkeys, that organisms with greater visual acuity had columns, suggesting that columns served a computational purpose. Recent findings from the tree shrew and squirrel, however, don't match the predictions of that hypothesis.
Additonally, the classic studies were all performed with extracellular electrodes, which sample neurons non-uniformly: neurons are identified by spiking, so highly-active neurons are more easily discovered, leading to their over-representation in experiments. In addition, recordings are easier and more stable in deeper layers, which generally correspond to thalamic input, rather than area-intrinsic computation.
More recent studies using two-photon calcium imaging, which works better on the superficial layers 2/3 and can detect weakly-active neurons, have generally not found columnar organization, or have found fuzzy, weak columns.
So today the reputation of the once-mighty column stands at a low point. Like other elements of the orthodoxy laid down by Hubel and Wiesel, further study with better tools has given us a more complicated picture than the tidy array of filters imagined 50 years ago (see the last few paragraphs on Hubel and Wiesel in this blog post for more on these critiques).
However, that doesn't mean these original findings are unimportant. They served as a major inspiration for decades of research on neurobiology, as well as for the highly successful "Hierarchical Temporal Memory" of Jeff Hawkins and Dileep George. The underlying principles, like vertical organization, anatomical maps, and receptive fields, are sound, and form the basis for quality neuroscientific research today.
Footnotes
† Subjects were anesthetized not with thiopental and succinylcholine, as Hubel and Wiesel did, but with Netflix and a vaporizer.
References
- Horton and Adams, 2005. "The cortical column: a structure without a function", Philos Trans R Soc Lond B Biol Sci.